Mitsis FJ. Hippocrates in the golden age: his life, his work and his contributions to dentistry. J Am Coll Dent. 1991; 58:26-30
Kassebaum NJ, Bernabe E, Dahiya M Global burden of severe periodontitis in 1990–2010: a systematic review and meta-regression. J Dent Res. 2014; 93:1045-1053 https://doi.org/10.1177/0022034514552491
Loe H, Anerud A, Boysen H, Morrison E. Natural history of periodontal disease in man. Rapid, moderate and no loss of attachment in Sri Lankan laborers 14 to 46 years of age. J Clin Periodontol. 1986; 13:431-445 https://doi.org/10.1111/j.1600-051x.1986.tb01487.x
Zhuang LF, Watt RM, Steiner S Subgingival microbiota of Sri Lankan tea labourers naive to oral hygiene measures. J Clin Periodontol. 2014; 41:433-441 https://doi.org/10.1111/jcpe.12230
Norderyd O, Hugoson A, Grusovin G. Risk of severe periodontal disease in a Swedish adult population. A longitudinal study. J Clin Periodontol. 1999; 26:608-615 https://doi.org/10.1034/j.1600-051x.1999.260908.x
Brinig MM, Lepp PW, Ouverney CC Prevalence of bacteria of division TM7 in human subgingival plaque and their association with disease. Appl Environ Microbiol. 2003; 69:1687-1694 https://doi.org/10.1128/AEM.69.3.1687-1694.2003
Ximenez-Fyvie LA, Haffajee AD, Socransky SS. Comparison of the microbiota of supra- and subgingival plaque in health and periodontitis. J Clin Periodontol. 2000; 27:648-657 https://doi.org/10.1034/j.1600-051x.2000.027009648.x
Kumar PS, Mason MR, Brooker MR, O'Brien K. Pyrosequencing reveals unique microbial signatures associated with healthy and failing dental implants. J Clin Periodontol. 2012; 39:425-433 https://doi.org/10.1111/j.1600-051X.2012.01856.x
Pelekos G, Ho SN, Acharya A A double-blind, paralleled-arm, placebo-controlled and randomized clinical trial of the effectiveness of probiotics as an adjunct in periodontal care. J Clin Periodontol. 2019; 46:1217-1227 https://doi.org/10.1111/jcpe.13191
Zheng H, Xu L, Wang Z Subgingival microbiome in patients with healthy and ailing dental implants. Sci Rep. 2015; 5 https://doi.org/10.1038/srep10948
Haubek D, Ennibi OK, Poulsen K Risk of aggressive periodontitis in adolescent carriers of the JP2 clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans in Morocco: a prospective longitudinal cohort study. Lancet. 2008; 371:237-242 https://doi.org/10.1016/S0140-6736(12)60135-X
Hajishengallis G. Periodontitis: from microbial immune subversion to systemic inflammation. Nat Rev Immunol. 2015; 15:30-44 https://doi.org/10.1038/nri3785
Van Dyke TE, Bartold PM, Reynolds EC. The nexus between periodontal inflammation and dysbiosis. Front Immunol. 2020; 11 https://doi.org/10.3389/fimmu.2020.00511
Lafaurie GI, Sabogal MA, Castillo DM Microbiome and microbial biofilm profiles of peri-implantitis: a systematic review. J Periodontol. 2017; 88:1066-1089 https://doi.org/10.1902/jop.2017.170123
Professor/Honorary Consultant in Periodontology; Centre for Host-Microbiome Interactions, Faculty of Dentistry, Oral and Craniofacial Sciences, King's College London
Throughout our lives, we continuously encounter micro-organisms that range from those essential for health, to those causing diseases. The oral cavity is permanently colonized by micro-organisms and our immune system has a challenging role in maintaining oral homeostasis by balancing its responses to this unique microbiota, while preserving the structural integrity of oral tissues. This article discusses relationships between oral microbial community structure and activity, specifically addressing the role of low-abundance microbial species and their interplay with the host immune system, in view of the pathogenesis of dental plaque-induced periodontal conditions.
CPD/Clinical Relevance: The knowledge of periodontal disease pathogenesis is critical for understanding periodontal risk assessment and designing effective, patient-centred treatment options.
Article
Unlike dental caries, which historically developed as a result of emerging civilizations, periodontal diseases are as old as humanity. They also affect wild and domesticated animals and continue to be one of the most prevalent diseases affecting humankind. Ancient Egyptians and the Chinese characterized periodontal diseases as inflammatory conditions, and Hippocrates discussed the aetiology, pathogenesis and treatment options for different forms of gum problems.1 Nowadays, severe periodontitis is the sixth-most prevalent condition worldwide, with an overall prevalence of 10.8% and around 743 million people affected.2 This figure corresponds surprisingly well with the seminal work of Loe et al. in Sri Lankan tea workers, who despite the fact of having high levels of plaque, calculus and gingival inflammation, showed rapid progression of periodontal disease in 8% of cases, while 11% of the examined population were largely free of alveolar bone loss.3 Modern, culture-independent methods have recently been used to characterize the subgingival microbiota of this ‘classical’ cohort of Sri Lankan tea workers, naïve to oral hygiene practices.4 Although significant differences in the overall subgingival microbial compositions were identified between shallow and deep periodontal sites, there was no obvious microbial clustering of the subjects according to their respective clinical progression groups, confirming the important role of the individual host response in the progression of the disease. Interestingly, improvements in plaque control and consequent reduction in mild disease in other cohort studies, did not necessarily result in similar reductions in the prevalence of severe periodontitis.5
Microbial species associated with periodontal diseases are part of the normal oral microbiota, but under certain ecological shifts, become pathobionts, acting never alone, but always as a community. It is not the mere presence of these micro-organisms that triggers the disease and drives its progression, but rather the interplay between divergent bacteria from different phyla and the host immune system.
Subgingival microbiota in periodontal health and disease
Microbial consortia in dental plaque have become the most characterized microbial communities in humans. Characterization of subgingival microbiota identified more than 500 species, some of which were found to be strongly associated with either clinically healthy or diseased periodontal sites. Periodontal diseases are polymicrobial infections, characterized by a microbial shift from Gram-positive to Gram-negative species. This microbial shift, found in diseased versus healthy sites, indicates that stability of the dental plaque communities is a good predictor of periodontal health, whereas changes in the composition of subgingival microbiota are associated with changes in the clinical status of periodontal tissues.6 The mechanisms that maintain the stability of, or induce changes in the microbial composition of, subgingival communities are just starting to emerge.
Studies evaluating the oral microbiome of clinically healthy individuals, employing 16S rRNA gene-sequencing, identified the predominance of Streptococcus, Haemophilus, Neisseria, Actinomyces and Rothia genera, which were present across ≥95% of samples. Interestingly, Gram-negative, anaerobic species traditionally associated with periodontal diseases, such as Porphyromonas gingivalis, Tannerella forsythia and Treponema denticola, have also been identified in the oral microbiome of persons with healthy gingival tissues, although at very low levels.7
The source of the newly formed subgingival microbiota around dental implants is not well understood. Saliva is a likely reservoir and conduit to transport micro-organisms from the rest of the dentition, and from non-dental sites, such as the tongue, buccal mucosa and tonsils, but simple geographic proximity is not a sufficient determinant for colonization of topographically distinct niches. Overall, dental implants consistently show lower microbial diversity in their subgingival environment compared to natural teeth. Healthy peri-implant crevices demonstrate higher levels of Prevotella, Treponema, Leptotrichia, Propionibacter and Lactococcus compared to teeth with healthy periodontium, but lower levels of Neisseria, Veillonella, Actinomyces and Fusobacterium species.8
In the absence of oral hygiene, the transition from periodontal health to gingivitis is accompanied by a shift in the bacterial community structure and increased diversity (Figure 1). Micro-organisms associated with gingivitis and bleeding on probing (BOP) include Campylobacter, Fusobacterium, Lautropia, Leptotrichia, Porphyromonas, Selenomonas, and Tannerella, while micro-organisms negatively correlated with BOP include Actinomyces, Rothia, and Streptococcus.
Figure 1.
(a, b) Scanning electron micrographs of subgingival dental plaque from a periodontitis patient. Courtesy of Dr Alexander Strachan, Plymouth Electron Microscopy Centre.
Rothia dentocariosa showed the most significant negative correlation with 1- and 2-week time points of experimental gingivitis, and with increased BOP scores.9 Nevertheless, the adjunctive use of probiotics, containing species associated with periodontal health, with non-surgical periodontal treatment, although positively affecting the composition of the oral microbiome, showed no additional clinical effectiveness when compared to non-surgical treatment alone.10
Despite the revolutionary advances that the invention and clinical application of dental implants have brought to oral rehabilitation, they also provide new, non-shedding, artificial surfaces prone to oral biofilm formation. Analogous to gingivitis, peri-implant mucositis is characterized by inflammation restricted to the peri-implant mucosa, without distinct alveolar bone loss. However, the periodontal microbiome demonstrates significantly higher diversity than the peri-implant, and distinct bacterial lineages are associated with health and disease in each ecosystem. From a microbial viewpoint, peri-implant mucositis appears to be a transitional phase on the course to peri-implantitis. Periodontal pathogens, such as Porphyromonas gingivalis, Tannerella forsythia, Prevotella intermedia and Capnocytophaga ochracea can be detected more frequently in peri-implant mucositis sites compared to gingivitis lesions.11
What does all of this mean to the dental practitioner?
Although distinct microbial communities are associated with periodontal/implant health and disease, it is not a clear-cut case. Stability of the subgingival microflora, as a whole, is more important than its individual constituents.
The host–pathogen interactions at subgingival niches
There is increasing evidence that the microbiome–immune system interplay is a major factor driving clinical phenotypes in the context of periodontal health and disease.
Pathogens, called generalists, are capable of infecting a wide range of hosts; while
Specialists establish an intimate relationship with only a particular group of genetically similar hosts.
Host tropism is the infection specificity of certain pathogens to particular hosts and host tissues. This type of tropism explains why some pathogens are only capable of infecting a limited range of host organisms. Aggregatibacter actinomycetemcomitans, a Gram-negative rod, which produces a leukotoxin that specifically lyses human neutrophils, displays significant genetic diversity. One particular clone, referred to as JP2, produces significantly enhanced levels of leukotoxin compared to the other lineages of this bacterium, which could, theoretically, lead to an enhanced potential to disrupt the immune defences of the periodontium. Population genetic analysis of A. actinomycetemcomitans strains from geographically dispersed individuals suggests that the JP2 clone originally emerged as a distinct genotype in the Mediterranean part of Africa over 2000 years ago, and subsequently spread to West Africa, from where it was transferred to North and South America. Interestingly, despite its now global dissemination, the JP2 clone still remains exclusively associated with individuals of West African descent indicating a strong host tropism effect.12
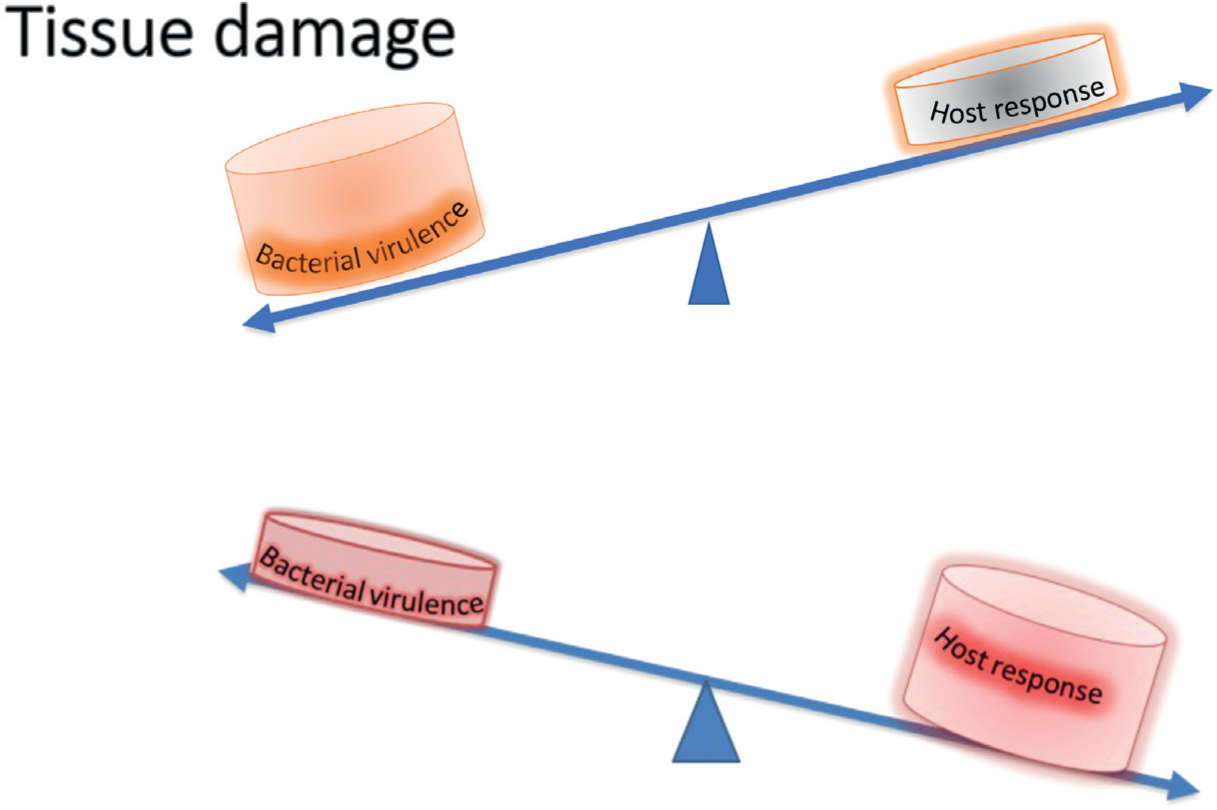
Periodontal inflammation begins as a protective response to subgingival biofilm. In susceptible individuals, this inflammation fails to resolve, and chronic inflammation becomes the periodontal pathology. Periodontal diseases result from excessive inflammation and may be considered a failure of inflammatory resolution pathways (Figure 2). Ageing is associated with the development of a low-level, systemic, chronic inflammation known as ‘inflammaging’. This chronic inflammatory state can contribute to diseases of ageing, one of which is periodontitis. The presence of inflammaging suggests a failure of the cell clearance mechanisms that ordinarily aid in the resolution of inflammation after pathogen infiltration or tissue injury. Cellular processes involved in the resolution of inflammation have been shown to become defective with age.13
Figure 2. Pathobiology of periodontal diseases. Tissue damage is caused by the direct action of microbial virulence factors or indirectly by excessive host-immune response.
What does all of this mean to the dental practitioner?
Genetic predisposition to particular periodontal pathogens and inability to tightly control inflammatory response could lead to increased susceptibility to periodontal diseases.
Pathogenesis of periodontal and peri-implant diseases
The microbial aetiology of periodontitis can be described as polymicrobial synergy and dysbiosis (PSD). The dysbiosis of the periodontal microbiota signifies a change in the relative abundance of individual components of the bacterial community compared to their abundance in health, leading to alterations in the host–microbial crosstalk, sufficient to mediate destructive inflammation and bone loss. Communication between the keystone periodontal pathogens, such as Porphyromonas gingivalis, and commensal micro-organisms, turns the latter into accessory pathogens, facilitates synergy between them and increases the pathogenicity of these dysbiotic subgingival communities. The central paradigm of this model is that the pathogenicity of the whole community is dictated by a subset of its constitutive species, and that this subgingival community of periodontal bacteria exhibits properties that are more than the sum of its constituent parts. Inflammation and dysbiosis positively reinforce each other since inflammatory tissue breakdown products are used as nutrients by the dysbiotic microbiota.14
Ecological studies have confirmed that periodontitis is a consequence of a dysbiotic shift in the microbiota driven by a change in the local environment. The microbiota of periodontal pockets gains an advantage from the substrates derived from inflammation and tissue breakdown, and this drives community restructuring. These findings have led to the newly proposed model of periodontal pathogenesis termed ‘inflammation-mediated polymicrobial-emergence and dysbiotic-exacerbation’.15
Although the clinical symptoms of peri-implantitis are similar to those of periodontitis, and both diseases are polymicrobial infections, the core microbiota of these diseases are different. The differences in topography and immunological characteristics of periodontal and peri-implant tissues drive to the logical conclusion that the biofilms associated with these surfaces are divergent. Peri-implantitis represents a heterogeneous, mixed infection that, beside periodontopathic micro-organisms, often includes uncultivable asaccharolytic anaerobic Gram-positive rods (Eubacterium and Filifactor spp), other anaerobic Gram-negative rods (Leptotrichia, Kingella, Mitsuokella spp), and in some cases opportunistic micro-organisms such as enteric rods and Staphylococcus aureus.16
What does all of this mean to the dental practitioner?
Interspecies communication in dental plaque, together with changes in the local environment (subgingival areas) may lead to establishment of pathogenic subgingival communities.
Clinical applications
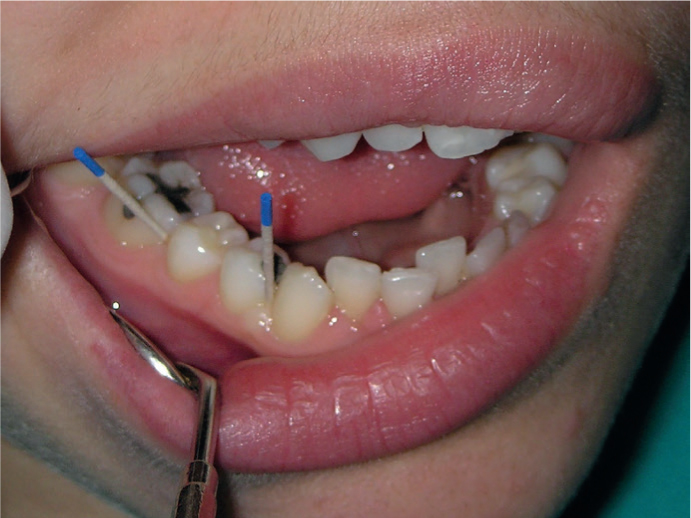
Alterations of normal human microbiome, or dysbiosis, can lead to diseases. In the future, personalized medicine may use microbiome-derived biomarkers for monitoring periodontal health. Disease progression may be more readily predicted and described by changes in microbiota than by clinical symptoms or the presence of certain pathogenic agents. Functional characterization of the oral microbiome may be more meaningful than identification of microbial community profiles due to functional redundancy among microbes (Figure 3).
Figure 3. Subgingival plaque sampling with paper points for site-specific periodontal treatment.
Better understanding of the complex mechanisms involved in the natural resolution of acute inflammation will allow us to exploit and develop novel therapeutic strategies to treat human inflammatory conditions, including periodontal diseases. The focus of future host–pathogen interactions research will be on how the body's natural healing pathways can be therapeutically harnessed for the treatment and prevention of periodontal disorders.